Introduction
Ephemeral pools, also commonly referred to as vernal pools, are short-term bodies of water that develop a complex ecosystem during periods of rainfall. Ephemeral pools support a variety of invertebrate organisms, with the class Branchiopoda, which includes fairy shrimp, clam shrimp and other invertebrates, being the most prominent (Huang et al., 2010). The differences in life found in these fragile ecosystems are dependent on the climate of the area as well as the type of rock of which the pools are made. These pools serve as a model of evolution as the communities are isolated and put through stresses that can promote selection of certain traits (Jocque et al., 2010). For example, due to the rapid evaporation rates in desert environments, evolution has favored the development of organisms that can complete their entire life cycle in periods as short as two weeks.
While many studies have focused on the animal life found in these environments, the microbial elements have been largely overlooked. Microorganisms fulfill various roles in ecosystems, ranging from symbionts to food sources, to providing various nutrients that are necessary for life in the ecosystem. The few microbiological studies that have been performed have hinted at a high diversity of bacteria that can vary considerably over even short distances (Altermatt & Ebert, 2010; Zeglin et al., 2011). The diversity of microorganisms at any particular site will likely be heavily influenced by the chemical composition of the water and the surrounding rock (Grathoff et al., 2007). In addition, as their habitat (the water) begins shrinking, increasingly intense competition may arise between the inhabitants of these pools as they compete for increasingly limited resources potentially causing major changes in the types and densities of the specific organisms that predominate over time (Wang et al., 2012). The purpose of this study was to characterize the bacteria and archaea found in six ephemeral pools located in southern Utah, and to identify a core set of microorganisms common to all these pools.
Methods
We studied six pools located within the Three Peaks Recreation Area (37°45’55.6"N 113°10’09.7"W) west of Cedar City, UT. We selected two pools from each of three locations situated approximately 1 km apart that had been previously observed to contain fairy shrimp (Order: Anostraca), which we used as an indicator of a healthy ecosystem. The ephemeral pools were hydrated with sterile reverse osmosis water to maintain constant water levels as water samples were collected every 96 hours for twelve days. Samples were submitted for microbiome analysis to the Microbial Analysis, Resources and Services (MARS) facility at the University of Connecticut for microbial community sequencing on the Illumina MiSeq system using Earth Microbiome Project 16S primers (Thompson et al., 2017) targeting the V4 region of bacterial and archaeal 16S rRNA-encoding genes.
We used the mothur bioinformatics analysis software (v1.44.3) to clean up and analyze the sequencing reads. Forward and reverse reads were combined and aligned with the SILVA reference database (Quast et al., 2013). Low-quality sequences and chimeras were removed with the VSEARCH algorithm. Duplicate sequences were merged, and taxonomic assignments were made using an 80% confidence cutoff with the Ribosomal Database Project reference database (Cole et al., 2014). The sequences were grouped into Operational Taxonomic Units (OTUs) with a 97% similarity threshold. We finally calculated the Alpha diversity (Shannon Diversity Index) of the microbes in the pools, determined the percent abundance of all taxa, and identified the genera common across all pools.
Results
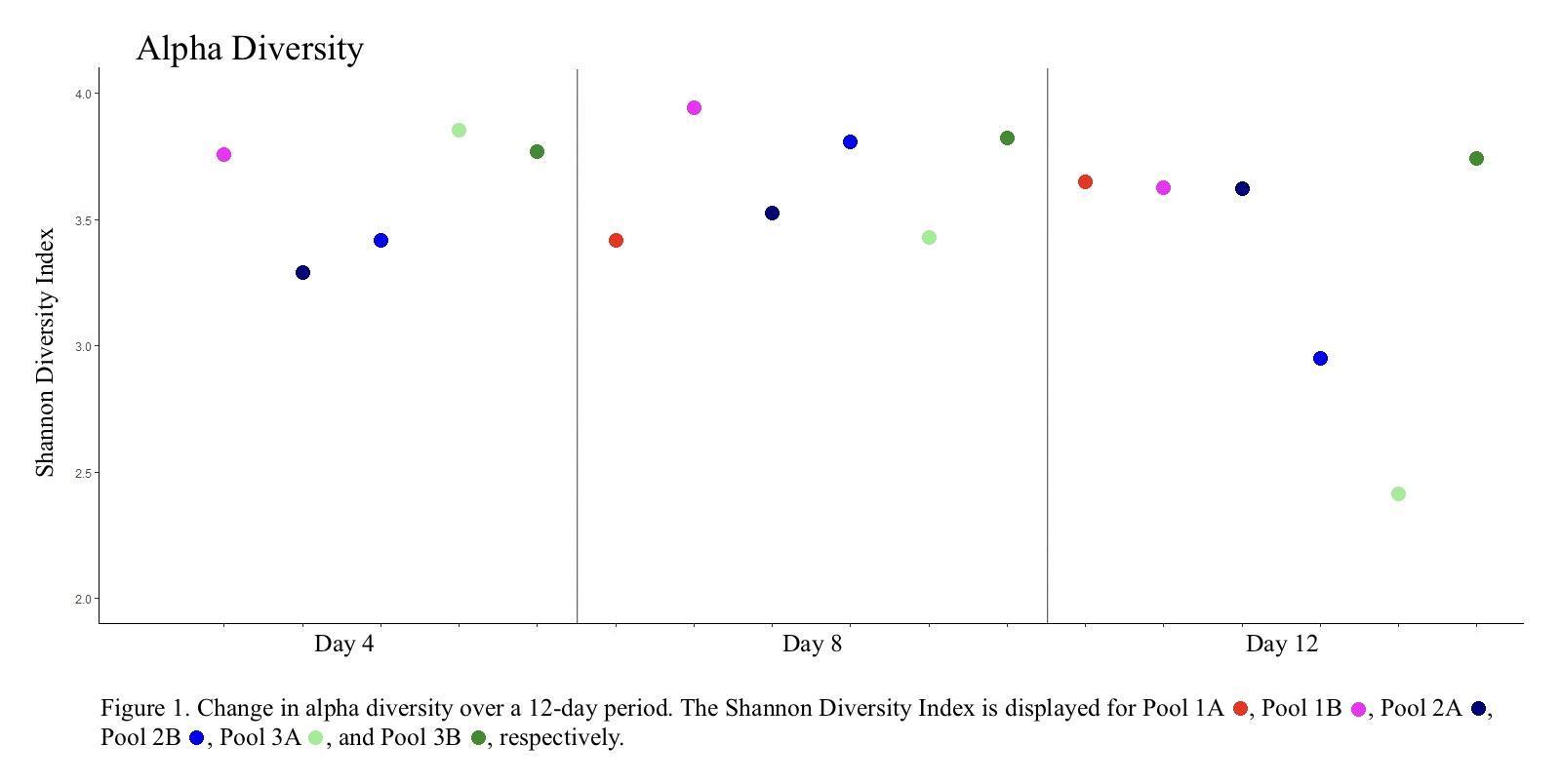
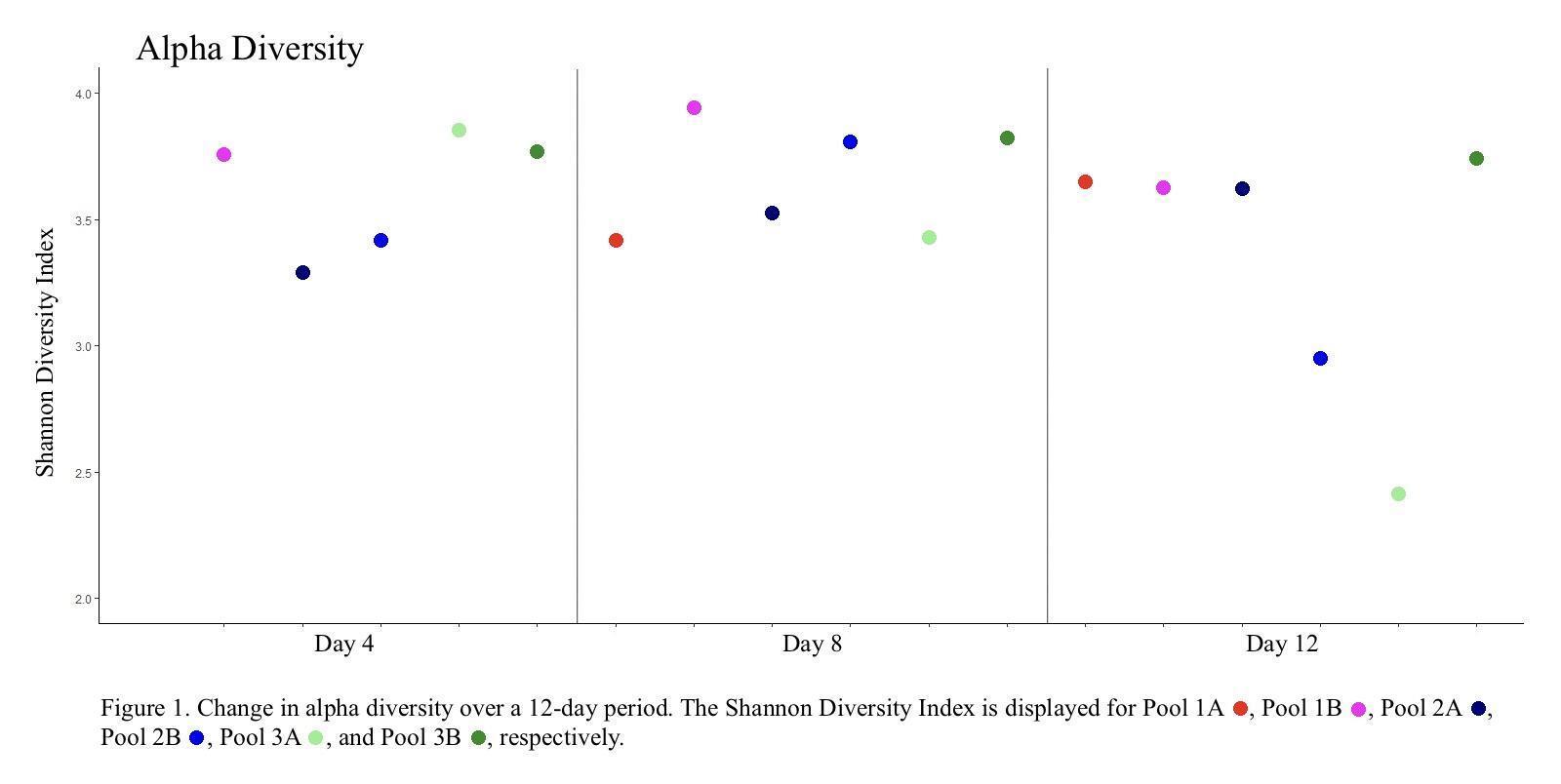
Except for two pools (Pool 2B and 3A) bacterial diversity remained relatively constant throughout the 12-day monitoring period with only minor fluctuations in diversity. Pools 2B and 3A, however, both experienced cyanobacterial blooms, with the percent abundance of cyanobacteria jumping from 1.4% and 2.9% to 42.2% and 58.6%, respectively (Day 4 to Day 12), and exhibiting a reduction in overall species diversity (Figure 1).
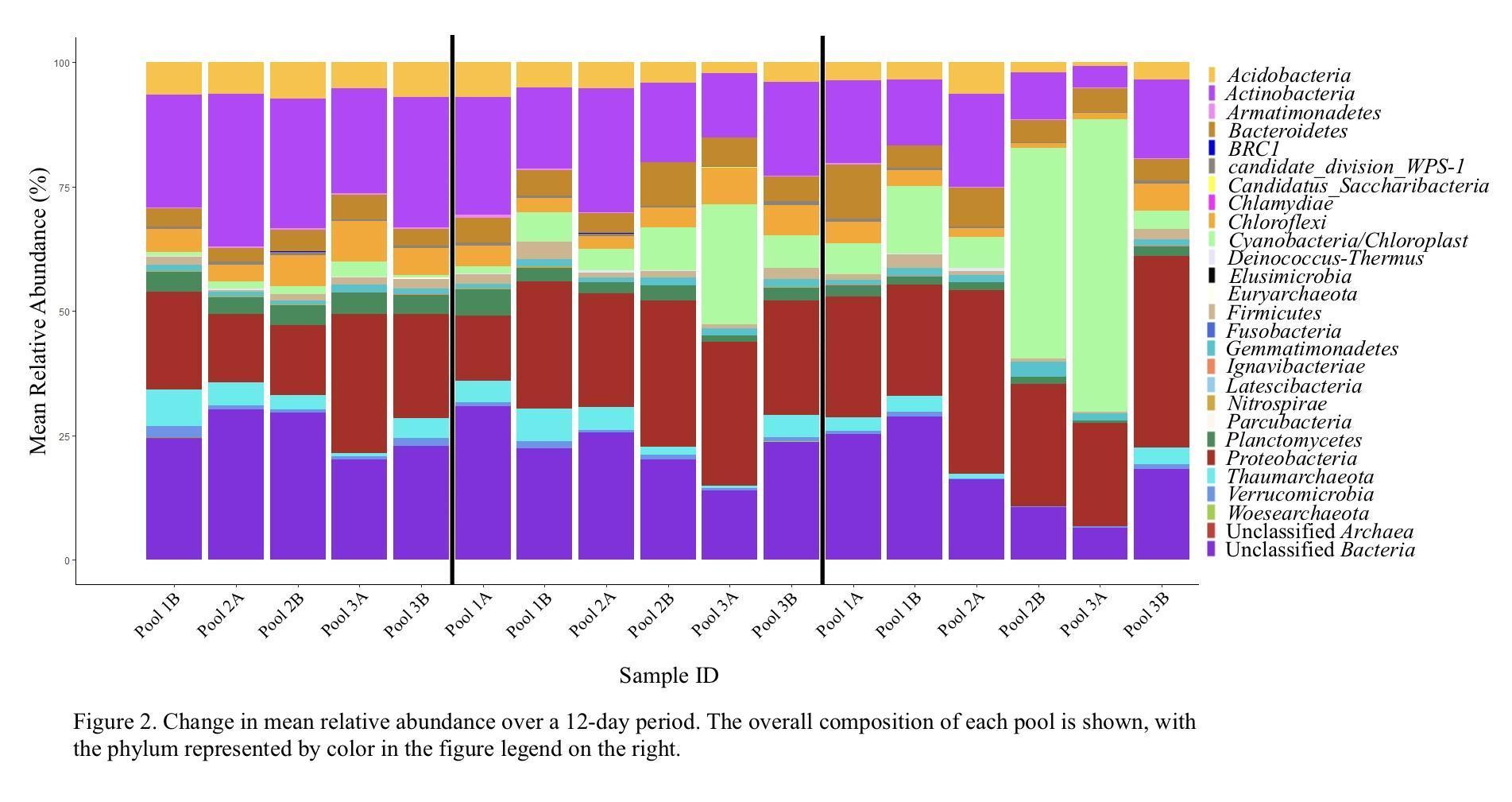
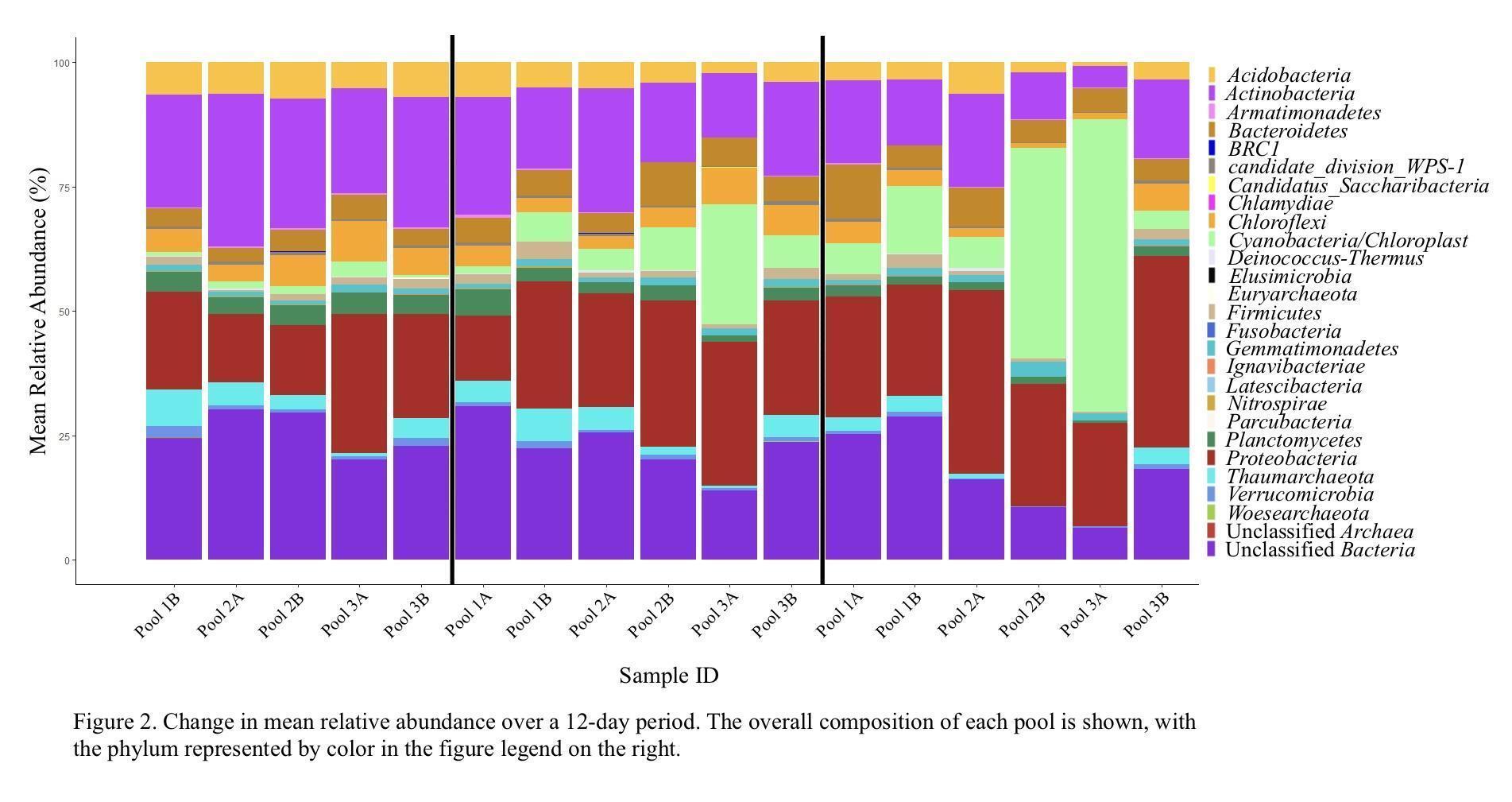
We observed a total of 23 bacterial and 4 archaeal phyla within these pools with 58 genera found to be common across the six pools, forming anywhere from 27.5% to 65.5% of the pool ecosystem. The percent average abundance for major phylum were observed as follows: Actinobacteria [18.7 ± 6.7%], Cyanobacteria [11.1 ± 16.2%], Proteobacteria [23.9 ± 7.1%], and Unclassified Bacteria [21.7 ± 7.0%] (Figure 2).
Discussion
A core of 58 OTUs were observed across all six ephemeral pools. Of these 25 OTUs were identified to the genus level and 33 consisted of unclassified bacteria. Many of the identified bacteria were common to soil and water environments, with some being plant symbionts and others known to break down different biochemical compounds in the environment, suggesting that these bacteria may play vital roles in the ephemeral pool ecosystem. As each site is located about 1 km apart, each pool is relatively isolated from the others, suggesting that with these 58 OTUs being found in common to all the sampled pools they may play important roles in these special ecosystems. Further research into the gut microbiome of the fairy shrimp found in these pools could elucidate connections between the bacteria and the fairy shrimp.
Two of the monitored pools (2B and 3A) experienced a cyanobacterial bloom resulting in a substantial decrease in alpha diversity. In a nitrogen-limited environment, nitrogen-fixing cyanobacteria would have a distinct advantage over other bacterial species (Huisman et al., 2018), and the subsequent rapid growth of cyanobacteria may have caused a drop in available nutrients, oxygen depletion, or the production of compounds that actively inhibited the growth of other bacterial and archaeal species. Interestingly, we observed that Proteobacteria and Bacteroidetes species were not as adversely impacted as other phyla, with the Proteobacteria increasing from 13.97% to 25.62% in Pool 2B and decreasing only slightly from 28% to 20.48% in Pool 3A. Likewise, the Bacteroidetes decreased only slightly, dropping from 5.0% to 4.63% in Pool 2B and from 4.70% to 4.56% in Pool 3A. Conversely, the mean relative abundance of other major phyla decreased 26.77% to 9.33% in Pool 2B and 20.62% to 3.84% in Pool 3A for the Actinobacteria, and from 29.23% to 10.28% in Pool 2B and 19.83% to 6.01% in Pool 3A for the Unclassified Bacteria. These patterns, specifically with Proteobacteria and Bacteroidetes being relatively unaffected, have been observed in other studies (Shao et al., 2014) and suggest that these bacteria generally continue to thrive when in close relation to cyanobacterial blooms. Likewise, previous studies have observed that Actinobacteria tend to be adversely affected by increases in Cyanobacteria and Proteobacteria populations (Ghai et al., 2014).
Our results identified Proteobacteria, Actinobacteria, and Acidobacteria to be major contributors to the ephemeral pool microbial community. These phyla are ubiquitous in soil and water ecosystems, primarily serving as decomposers (Carrino-Kyker & Swanson, 2008; Ghai et al., 2014; Montiel-Molina et al., 2021, and Ibekwe et al. 2006), supporting the observation that ephemeral pools are generally heterotrophic environments (Carrino-Kyker & Swanson, 2008). Further study of the specific genera of these bacteria will help illuminate their environmental niche in these unique environments,
Our study has provided valuable groundwork in not only understanding the composition of these fragile ecosystems, but in providing insight into the microbial component of potential conservation efforts. A set of 58 OTUs was found to be common across the six ephemeral pools monitored in this study. Of these 58 OTUs, 33 (57%) consisted of unidentified bacterial genera, which may potentially play novel functional roles and/or form interactions crucial to the health of these unique ecosystems. With climate change threatening to alter precipitation and temperatures, tenuous ecosystems, including ephemeral pools and the flora and fauna they support, could be severely impacted (Franksen et al. 2022). Within our study we observed an example of ecological disruption when cyanobacterial blooms in two of the pools resulted in a severe decrease in overall microbial diversity. Any imbalance has the potential to disrupt crucial interactions within an ecosystem resulting in devastating declines in species richness and in the ability of the ecosystem to survive. This study focused on the microorganisms present in ephemeral pools but it must be noted that a variety of invertebrates and vertebrates are also present in these ecosystems and that disruptions at the microscopic level have a potential impact on all levels of a trophic system. To fully understand an environment and the life it contains, all interactions must be considered, including microbe-microbe and host-microbe interactions (Amato, 2013). By providing insights into the environmental and microbiotic elements of an ecosystem, such as our study has done with ephemeral pools, conservation efforts for the flora and fauna found within and supported by them can be enhanced.
Acknowledgements
We would like to give special thanks to Emily Burns for her assistance in generating graphs via R-Studio, as well as assistance in data processing. We would also like to acknowledge Claire Gehl and Asia Negron in their help with sample collection. This study was performed under a Memorandum of Understanding (MOU) and Data Sharing Agreement (DSA) with the United States Department of the Interior, Bureau of Land Management (BLM), Cedar City Field office; we are grateful for the support of this office in allowing us to conduct research on lands managed by the BLM.