Introduction
Most animal species are known to have a variety of food choices depending on their physical state (Huffman, 2003). Insects also exhibit signs of feeding behavior change with response to stressors (McNamara, 2011). Cabbage looper moths (Trichoplusia ni) are herbivores that typically feed on lower growing vegetation, specifically cultivated plants and weeds. Cabbage loopers are known for having a specific taste in species of plant in the larvae stage and then expanding their diet to a wider variety by the fourth or fifth larvae (Schoonhoven, 1987). They are notorious for causing severe damage to crop and gardens because they are very heavy feeders leading to most larvae consuming up to three times its weight in plant material daily. The variety of species they consume can vary greatly. This variance in diet with age progression and feeding preference can have a serious trophic impact on native and non-native plant competition in a given area (McNamara, 2011). One of their main stressors are general parasitoid wasps. The female wasp uses the caterpillar as a host for its eggs (studies suggesting 2nd instar caterpillar preference). Once injected the eggs have a safe place to mature as the caterpillar continues to grow and act as a protector for the eggs (Singer et al., 2009). Anywhere from 40-80 eggs will emerge from the body of the caterpillar to pupate, still attached to the host. Once hatched the eggs overtake the caterpillar feeding on the internal organs and body of the caterpillar, resulting in host termination (Price, 1973).
Different phytochemically active plants can be a large determinant of the effectivity of immune response from the caterpillars (Singer et al., 2009). As they grow, and their tolerance adapts and expands, they are introduced to a higher variety of food sources, allowing them to interact and experience different nutrients and chemicals within the plants they consume. Providing a type of chemical buffet in which the caterpillars can pull from depending on what their situation. Instead of choosing plants that they normally feed on, some caterpillars choose plants with different active phytochemicals that turn out to be harmful for the caterpillar (Huffman, 2003). One of and the most supported hypothesis is that the caterpillar is trying trigger greater immune responses by changing their diet to different phytochemically active plants (Singer et al., 2009). Knowing that the caterpillars are opting to change their diet to a less favorable species of plant, suggests that this is conscious behavioral change to ward off the parasitoid (like self-medication) (Huffman, 2003). Smilinach (2009) conducted a study with a similar species, in which they found that the immune response is clearly one of the most effective defenses that caterpillars have against parasitism (Smilanich et al., 2009). Smilinach (2011) looked at specific phytochemicals and their effect on behavior, showing results that when allowed to choose among three host plant species, individuals harboring early-stage parasitoids increased their consumption of a nutritious plant containing antioxidants. Our hypothesis is that the cabbage loopers percent consumed will vary from that of the control group depending on the injection type, showing data suggesting a feeding behavior change in relation to interaction with a parasitoid.
Methods
I used two different plant species, Mimulus guttatus and Nasturtium officinale. These two plants vary in phytochemical activity and also compete in the wild. Garden Nasturtium officinale is a good source of micro elements such as potassium, phosphorus, calcium, and magnesium, along with macro elements like zinc, copper, and iron (Huffman, 2003). Mimulus guttatus is far less chemically active and is more of a pollinator plant.
I ordered caterpillars around 4-5th instar and let them feed on leaves from one specific nasturtium or Mimulus guttatus individual. Each caterpillar was assigned an individual and allowed to feed on only that individual for the remainder of the study. With a sample size n=65 caterpillars, we assigned a group of caterpillars to Nasturtium officinale (30) and a group to the monkeyflower (30) and a group to artificial diet (5). We also assigned groups to both plants that will be stabbed and injected SI (26), a group stabbed and not injected SN (26) a group to both plants that will not be tampered with at all N (13). A group was included that is stabbed and not injection to see if there is a behavior change purely from the interaction with the wasp and not the actual egg implantation. The injection process occurred on the same day, with as little time between start and finish to avoid other variables. Using silica beads (around 40 micrometers in diameter), dyed red for higher rate of successful injection, we injected into the back of the caterpillars just behind the front legs, mimicking the wasp egg injection effect using glass insulin needles. After injection, we replaced all leaves, taking photos before and after a 24-hour feeding period. That 24-hour period will conclude the trials. After the 24-hour period the caterpillars will be freeze killed. Using ImageJ, we compared all the photos to see if there was a difference in amount consumed among injection and non-injection groups to see what eating behavior changes occurred. All plant species were grown at the Utah Valley University Greenhouse in Orem Utah. 60 different individuals each grown as clippings from different populations collected throughout North America, including Nasturtium officinale populations that are both cultivated and natural. Work has been done in the lab to show the different competition results of these two species with no outside stressors, directly in the greenhouse that will eventually be as a good project after results are collected from this project to see the effects herbivores have on competition success rates. We ran an ANOVA to find the difference between interaction type and percent tissue consumption. All statistics were completed in R V. 3.5.
Results
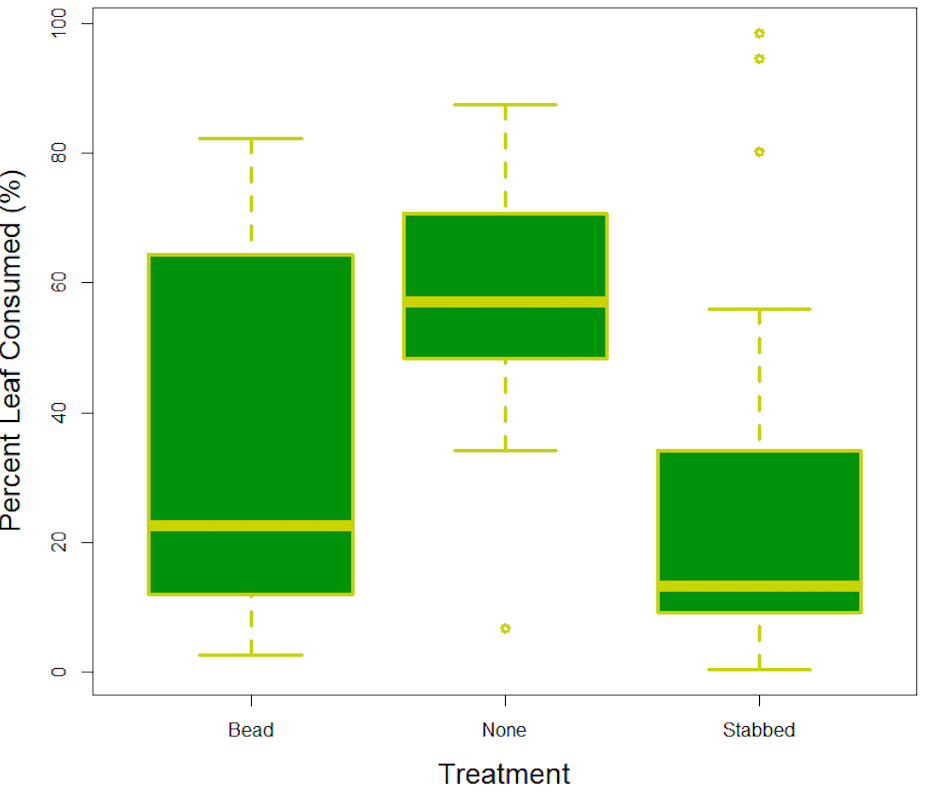
For percent of leaf consumed by Injection type was significantly different (F2,49 = 3.909, p = 0.026). Caterpillars that were stabbed but not injected were significantly different than those that were not stabbed (p=0.021). Plant type and plant type with an interaction term of injection type were not significantly different in percent of leaf tissue consumed (F1,49 = 0.562, p = 0.45 and F2,49 = 1.136, p = 0.32 respectively). This data suggests a clear difference in appetite with respect to interaction. With the group that was stabbed and not injected having the most significant difference in appetite with the lowest average percent consumed. The Stabbed and Injected average was still lower than the control group, suggesting a lowering of percent consumed after being interacted with at some capacity. There was no difference between percent consumed and plant type, which was not something we expected, however more of a difference between percent consumed and interaction type representing a feeding behavior change that was significant.

Discussion
This project showed the different feeding behavior changes in cabbage loopers in response to parasitoids. Our results suggested with regard to percent of leaf consumed, Injection type was not significantly different, but caterpillars that were stabbed without injection were significantly different than those that were not stabbed. This is likely due to the process of defensive evolution that has occurred in many plant species over time, in response to being attacked by herbivorous insects. Many species of plant have developed chemical defenses composed of different compounds that provide various defensive attributes, for example giving the plant a bitter taste or containing glycosides that react with enzymes to cut off cellular respiration (McNamara, 2011). I think this is how herbivores can differentiate between different phytochemically active plants. Our hypothesis that percent consumed averages would change significantly based on injection was half right. At least suggesting some behavior change relative to tissue consumption and injection type.
In the future our research will look at the immune response triggered by the different phytochemically active plants. Using the red dyed silica beads that we injected, we can dissect and measure total encapsulation (melanization effect around the bead) to score immune response (Smilanich et al., 2011). We can use that data to compare plant types with regard to immune response effectivity, as well as compare tissue consumption to immune response scores. This way we can begin to identify why and how the cabbage loopers have the behavior change we see in this study. The result that cabbage loopers are significantly changing feeding behaviors, we can also discuss the difference in preference with the presence of more than one leaf. In this study, our aim was to find data that suggests the significance between injection type and feeding behavior change by tissue consumption on a given leaf. Now, we can look at injection type with tissue consumption with more than one given leaf, which might give us insight on whether there is more than just appetite change, and instead a greater selectivity among the diet choice.
Cabbage loopers are an important component of terrestrial food systems and their ability to consume or be consumed in a system can dictate bottom-up and top-down trophic ecosystem structures. By not feeding on the margin and piercing large holes in leaves they have a serious effect on plant mortality. Based on diet preference they can have an effect on crop yield, native/non-native plant competition, and other environmental effects (Capinera, 1999).