Introduction
Sympatric speciation has been relatively little studied. This may be due to the inability of researchers to find clear systems and models of sympatric speciation as well as the, sometimes, controversial issues surrounding this mode of evolution. Sympatric speciation is the process of a population diverging within a close geographic area to produce two new species. This process is dependent on the breaking of gene flow within a widely interbreeding population and the subsequent development of phenotypic variation occurs (Les et al., 2015).
There are many factors that contribute to speciation events. Importantly in plants, the loss of a flow of pollen may lead to speciation (Butlin, 1987). Pollinators play a role in pollen flow and plant sexual traits most likely contribute to why pollinators may prefer some plants over others (Urbanowicz et al., 2020). Tied to these pollinators is the phenological (or timing) of events in the plant’s life. Plants may not be able to mate and maintain gene flow if they miss each other. The difference in flowering time, starting their lives, and growth can all force pollinators to not have the opportunity to mate (Sianta & Kay, 2021).
One important difference in many plants is the success of their seeds. Seed germination and days till germination can vary widely between plant species. These factors may cause speciation within clusters of sympatrically growing seed plants (Willis et al., 2014). Some plants will put large energy resources into their seeds, allowing for a greater germination percent of their seeds. On the other hand, some plants can create more seeds with lower germination percentages and succeed in spreading many propagules with a small but acceptable germination rate. The timing of the germination event can also play a key role in ecological separation as a delay of several weeks may cause plants with differing germination to develop differently. Consequentially separating their flowering dates and isolating them sexually leads to the maintenance of genetic differentiation (Daws et al., 2002).
In this study, we used the common western species of rabbitbrush (Ericameria nauseosa) to test the ecological segregation of several broad subspecies. We predicted that these different subspecies groups would differ in total percent germination and the days till germination. Thus, reflecting shifts in phenotypes leading to an ongoing speciation event within rabbitbrush.
Methods
Rabbitbrush (Ericameria nauseosa) is a native perennial shrub in the Asteraceae family. It has erect stems and commonly has numerous leaves. When flowers bloom in late summer and fall they are bright yellow and are in clusters. It thrives in pinyon-juniper areas, dry areas, and rocky mountainsides at low elevations. Around 21 species of rabbitbrush have been described within North America and placed in gray or green-colored groups, both of which can be deciduous or non-deciduous. Within these subspecies, ranges are variable with some being very narrowly isolated, but many being widely distributed with overlapping ranges (McArthur & Welch, 1986). Within these overlapping subspecies they are often found growing together. These subspecies have generally been defined by phenotypic variation that is often hard to distinguish in the field. However, other studies have shown the importance of the phytochemical makeup of these species as well as the predilection of galling insects for specific subspecies (Floate et al., 1996). Despite these morphological and ecological differences little if any work has been done on the ability of pollinators to discriminate between subspecies, resulting in a significant gap in our knowledge of how these subspecies are maintained.
To test this hypothesis of pollinator discrimination of subspecies of rabbitbrush we started with a germination experiment focusing on total percent germination and the days till germination concentrating on color groups and deciduousness.
Using previously collected seeds from ten locations in Utah, Salt Lake, and Duchesne Counties in Utah during 2021, each seed was identified as being in the green or gray group as well as if it was deciduous or not. Ten individual seeds from each collected specimen were planted in 80% peat 20% perlite all-purpose soil. Each seed was planted in an individual cell in a plug tray and dusted with 80% peat and 20% perlite germination soil. All the plants were grown in a greenhouse with consistent watering, light, and temperature at 20°C - 27°C (Meyer et al., 1989), and fertilizer treatments once a week (1:100 ppm with 20-10-20 fertilizer). The seeds were observed consistently over 3 months and the specific date of germination was recorded. Specimens were confirmed using keys from, “Proceedings– Symposium on the Biology of Artemisia and Chrysothamnus” (McArthur & Welch, 1986). The specimens were also confirmed by local botanists. Additionally, voucher specimens of the plants were collected and deposited into the Utah Valley University herbarium (UVSC).
To look for differences between subspecies groups we used a two-way ANOVA. We also included an interaction term in order to test for the interactions of the deciduous and color groups. All statistical analysis was done in R.
Results
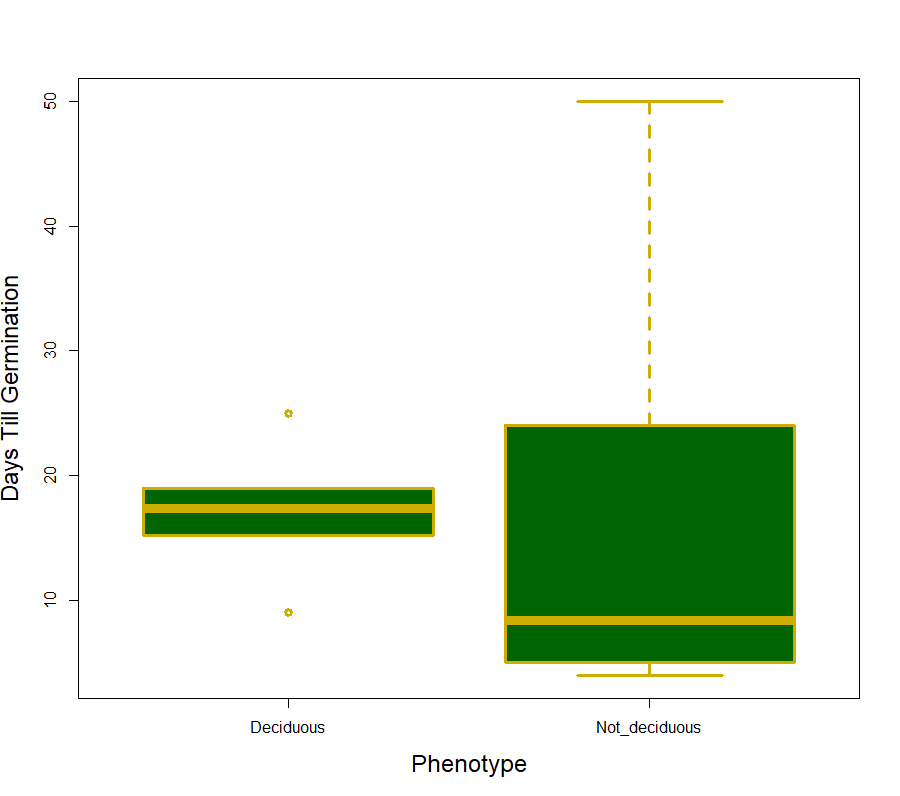
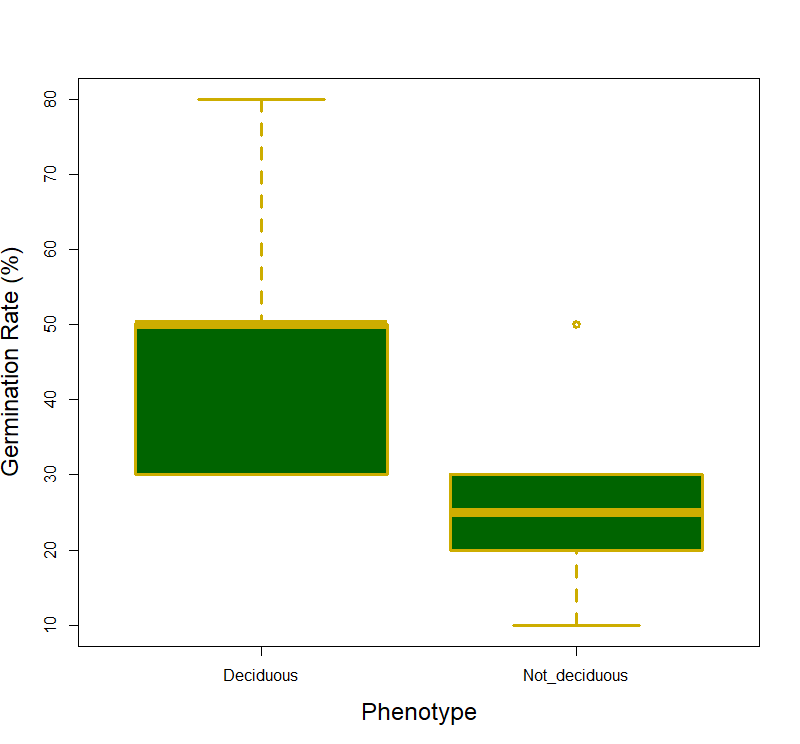
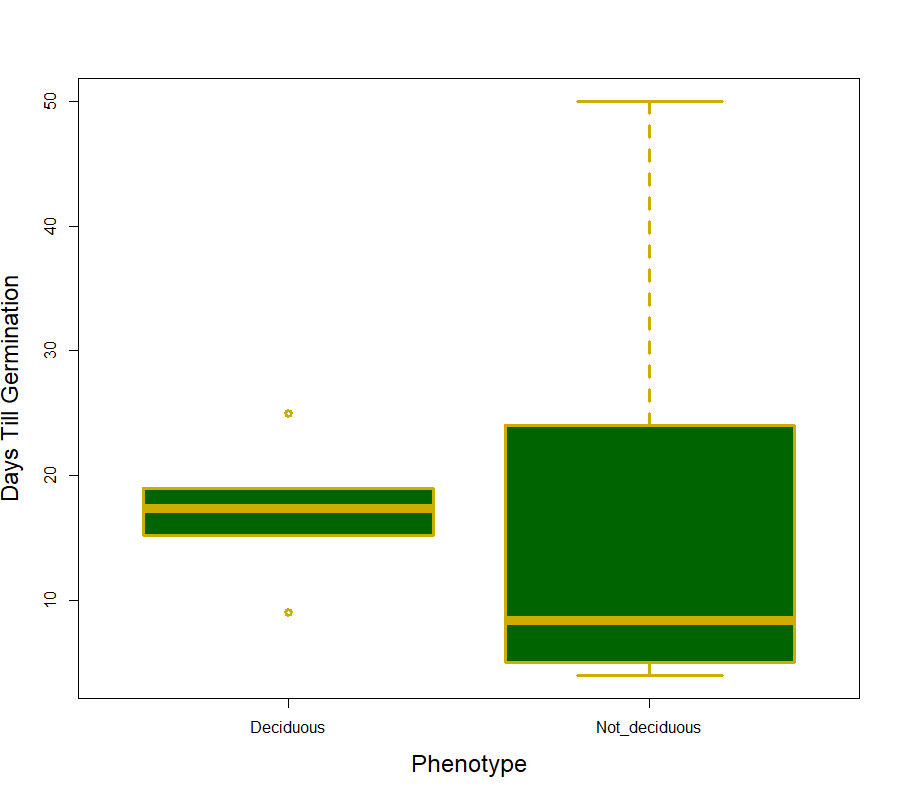
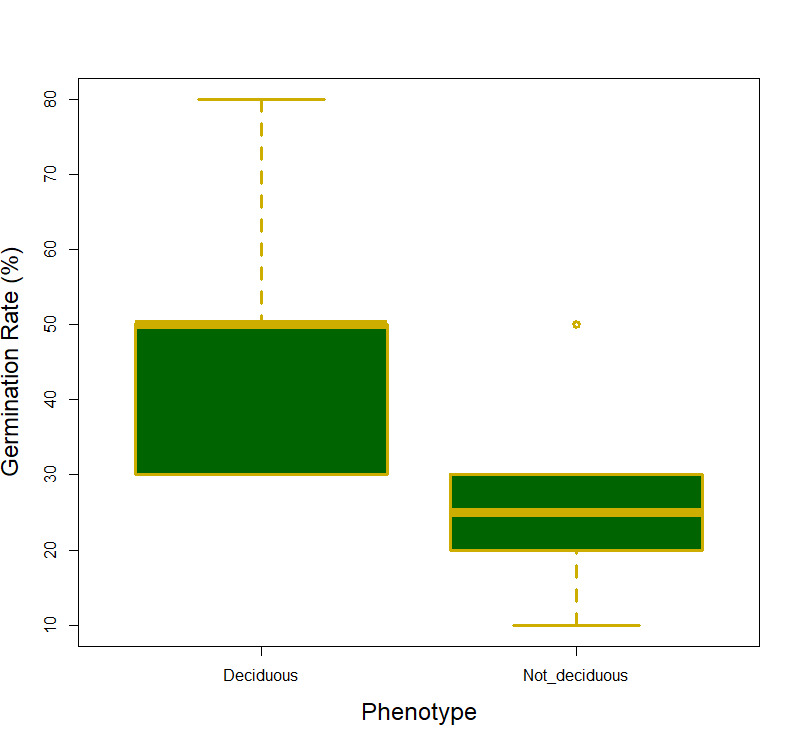
We did not support our hypothesis that germination percent and days till germination would differ between different color phenotypes (F1,14 = 2.31, p =0.15 and F1,14 = 0.634, p =0.439 respectively). When it came to differences between the deciduous phenotypes we did not find a significant difference in days till germination (F1,10 = 0.005, p =0.946; Fig 1), however, the deciduous phenotypes did differ in germination percent with the deciduous phenotype having a significantly large percentage of seeds germinating (F1,10 = 5.38, p =0.048; Fig 2). The interaction term (deciduous groups and color groups) was not significant for any germination variable (F1,8 = 1.29, p =0.300).
Discussion
Though our hypothesis was not supported by our results concerning germination percent and days till germination in regards to the gray and green phenotypes, we did see a significance in germination percent and if the plant was deciduous or not. These differences in seed germination percent tell us that there is an importance about that relationship in rabbitbrush. This information leads to a hopeful understanding of how speciation events may be occurring between different subspecies of rabbitbrush with respect to the deciduous phenotype.
The insignificant results most likely show a considerable niche overlap between the subspecies. Although the two phenotypes have unique genetically based traits, gene flow is likely active between them and thereby not experiencing much divergence. Additionally, these traits may not be under selective pressure whereas sexual traits (like pollination) may be a more suitable characteristic to consider when it comes to potential ongoing speciation events within the subspecies (Rundle & Nosil, 2005).
There was one significant result concerning the germination percentage and whether the subspecies were deciduous or not. This may be due to the non-deciduous plants being at a disadvantage by putting resources into forming seeds and regrowing leaves yearly. For example, many evergreen plants in deciduous forests employ tradeoffs in seed germination for chances to increase primary production and live longer lives (Nakashizuka, 2001). This tradeoff could be beneficial in some systems but probably not in a controlled environment in our greenhouse.
In the grand scheme of things, these results contribute to our next step in an additional study on pollinator discrimination of subspecies. Specifically, the genetic makeup and morphological differences between the subspecies of rabbitbrush may be a reason that speciation events occur among rabbitbrush (Baack & Stanton, 2005). In addition, we are considering the factor that pollinator discrimination of floral traits may also contribute to these speciation events (Zhao & Huang, 2013). We will be observing pollinator communities and pollen flow among the different subspecies and comparing and contrasting floral morphological measurements.